Describe What Happens to the Chromosomes When an Egg Cell and Sperm Cell Join.
Once released, egg and sperm alike are destined to die inside minutes or hours unless they find each other and fuse in the process of fertilization. Through fertilization, the egg and sperm are saved: the egg is activated to brainstorm its developmental program, and the haploid nuclei of the two gametes come together to course the genome of a new diploid organism. The mechanism of fertilization has been most intensively studied in marine invertebrates, peculiarly sea urchins. In these organisms fertilization occurs in sea water, into which huge numbers of both sperm and eggs are released. Such external fertilization has been more accessible to study than the internal fertilization of mammals, which normally occurs in the female reproductive tract subsequently mating.
In the late 1950s, however, it became possible to fertilize mammalian eggs in vitro, opening the way to an analysis of the cellular and molecular events in mammalian fertilization. Progress in agreement mammalian fertilization has brought substantial medical benefit: mammalian eggs that take been fertilized in vitro tin develop into normal individuals when transplanted into the uterus; in this way many previously infertile women have been able to produce normal children. As mentioned earlier, it is possible to use in vitro fertilization to produce a clone of a sheep, a squealer, or a mouse past transferring the nucleus of one of its somatic cells into an unfertilized egg that has had its own nucleus removed or destroyed. At that place is no reason to dubiousness that a man could be cloned in the same way, although there are serious upstanding arguments about whether this should ever be done, especially as the likelihood of producing an abnormal child is very loftier. In this section, we focus our give-and-take on the fertilization of mammalian eggs.
Species-Specific Binding to the Zona Pellucida Induces the Sperm to Undergo an Acrosome Reaction
Of the 300,000,000 human being sperm ejaculated during coitus, only about 200 reach the site of fertilization in the oviduct. There is evidence that chemical signals released by the follicle cells that surround the ovulated egg concenter the sperm to the egg, merely the nature of the chemoattractant molecules is unknown. In one case information technology finds an egg, the sperm must first migrate through the layer of follicle cells and and then bind to and cantankerous the egg glaze—the zona pellucida. Finally, the sperm must bind to and fuse with the egg plasma membrane. To become competent to attain these tasks, ejaculated mammalian sperm must normally be modified by weather condition in the female reproductive tract, a process called capacitation, which requires about v–6 hours in humans. Capacitation is triggered by bicarbonate ions (HCO3–) in the vagina, which enter the sperm and straight actuate a soluble adenylyl cyclase enzyme in the cytosol. The cyclase produces cyclic AMP (discussed in Chapter xv), which helps to initiate the changes associated with capacitation. Capacitation alters the lipid and glycoprotein composition of the sperm plasma membrane, increases sperm metabolism and motility, and markedly decreases the membrane potential (that is, the membrane potential moves to a more negative value so that the membrane becomes hyperpolarized).
Once a capacitated sperm has penetrated the layer of follicle cells, information technology binds to the zona pellucida (run into Figure 20-21). The zona unremarkably acts as a bulwark to fertilization across species, and removing information technology frequently eliminates this barrier. Human sperm, for example, volition fertilize hamster eggs from which the zona has been removed with specific enzymes; not surprisingly, such hybrid zygotes fail to develop. Zona-free hamster eggs, however, are sometimes used in infertility clinics to assess the fertilizing capacity of human sperm in vitro (Effigy 20-30).

Figure 20-thirty
Scanning electron micrograph of a human sperm contacting a hamster egg. The zona pellucida of the egg has been removed, exposing the plasma membrane, which contains numerous microvilli. The ability of an private's sperm to penetrate hamster eggs is (more...)
The zona pellucida of mammalian eggs is composed mainly of 3 glycoproteins, all of which are produced exclusively past the growing oocyte. 2 of them, ZP2 and ZP3, gather into long filaments, while the other, ZP1, cross-links the filaments into a three-dimensional network. The protein ZP3 is crucial: female mice with an inactivated ZP3 cistron produce eggs lacking a zona and are infertile. ZP3 is responsible for the species-specific binding of sperm to the zona, at least in mice. Several proteins on the sperm surface that bind to specific O-linked oligosaccharides on ZP3 take been implicated equally ZP3 receptors, simply the contribution of each is uncertain. On bounden to the zona, the sperm is induced to undergo the acrosome reaction, in which the contents of the acrosome are released by exocytosis (Figure xx-31). In the mouse, at least, the trigger for the acrosome reaction is ZP3 in the zona, which induces an influx of Ca2+ into the sperm cytosol; this in turn initiates exocytosis. An increase in cytosolic Ca2+ seems to be necessary and sufficient to trigger the acrosome reaction in all animals.
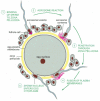
Figure 20-31
The acrosome reaction that occurs when a mammalian sperm fertilizes an egg. In mice, a unmarried glycoprotein in the zona pellucida, ZP3, is idea to be responsible for both binding the sperm and inducing the acrosome reaction. Note that a mammalian sperm (more...)
The acrosome reaction is required for fertilization. Information technology exposes various hydrolytic enzymes that assist the sperm tunnel through the zona pellucida, and it exposes other proteins on the sperm surface that bind to the ZP2 poly peptide and thereby help the sperm maintain its tight binding to the zona while burrowing through it. In addition, the acrosome reaction exposes proteins in the sperm plasma membrane that mediate the bounden and fusion of this membrane with that of the egg, as we hash out beneath. Although fertilization normally occurs past sperm—egg fusion, information technology can too be accomplished artificially, past injecting the sperm into the egg cytoplasm; this is sometimes done in infertility clinics when at that place is a problem with sperm—egg fusion.
The Egg Cortical Reaction Helps to Ensure That Just One Sperm Fertilizes the Egg
Although many sperm can bind to an egg, ordinarily only i fuses with the egg plasma membrane and injects its nucleus and other organelles into the egg cytoplasm. If more than one sperm fuses—a condition chosen polyspermy—multipolar or extra mitotic spindles are formed, resulting in faulty segregation of chromosomes during cell segmentation; nondiploid cells are produced, and development commonly stops. Two mechanisms can operate to ensure that only one sperm fertilizes the egg. In many cases, a rapid depolarization of the egg plasma membrane, which is caused past the fusion of the kickoff sperm, prevents further sperm from fusing and thereby acts as a fast primary block to polyspermy. But the membrane potential returns to normal shortly after fertilization, so that a second mechanism is required to ensure a longer-term, secondary block to polyspermy. This is provided by the egg cortical reaction.
When the sperm fuses with the egg plasma membrane, it causes a local increase in cytosolic Ca2+, which spreads through the prison cell in a moving ridge. In some mammalian eggs, the initial increase in Ca2+ is followed by prolonged Ca2+ oscillations. There is testify that the Catwo+ wave or oscillations are induced by a protein that is introduced into the egg by the sperm, simply the nature of the protein is unknown.
The Caii+ moving ridge or oscillations actuate the egg to begin development, and they initiate the cortical reaction, in which the cortical granules release their contents past exocytosis. If the cytosolic concentration of Ca2+ is increased artificially—either directly by an injection of Ca2+ or indirectly by the use of Ca2+-conveying ionophores (discussed in Affiliate eleven)—the eggs of all animals then far tested, including mammals, are activated. Conversely, preventing the increment in Ca2+ by injecting the Caii+ chelator EGTA inhibits activation of the egg in response to fertilization. The contents of the cortical granules include various enzymes that are released by the cortical reaction and alter the construction of the zona pellucida. The altered zona becomes "hardened," and so that sperm no longer bind to it, and it therefore provides a block to polyspermy. Amidst the changes that occur in the zona is the proteolytic cleavage of ZP2 and the hydrolysis of sugar groups on ZP3 (Effigy 20-32).
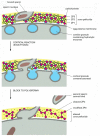
Figure twenty-32
How the cortical reaction in a mouse egg is thought to foreclose additional sperm from entering the egg. The released contents of the cortical granules both remove carbohydrate from ZP3 so it no longer can bind to the sperm plasma membrane and partly cleave (more...)
The Machinery of Sperm—Egg Fusion Is Still Unknown
After a sperm has penetrated the extracellular coat of the egg, information technology interacts with the egg plasma membrane overlying the tips of microvilli on the egg surface (see Figure 20-30). Neighboring microvilli then rapidly elongate and cluster around the sperm to ensure that it is held firmly so that it can fuse with the egg. Subsequently fusion, the entire sperm is drawn caput-get-go into the egg as the microvilli are resorbed. In mouse sperm, a transmembrane protein called fertilin, which becomes exposed on the sperm surface during the acrosome reaction, helps the sperm bind to the egg plasma membrane and may as well accept a part in the fusion of the two plasma membranes.
Fertilin is composed of two glycosylated transmembrane subunits called α and β, which are held together past noncovalent bonds (Effigy 20-33). The extracellular N-terminal domain of the fertilin subunits is thought to demark to integrins in the egg plasma membrane and thereby assist the sperm adhere to the egg membrane in grooming for fusion. The integrin in the egg plasma membrane is associated with a fellow member of the tetraspan family of membrane proteins—so-called because they accept iv membrane-spanning segments. Female mice that are scarce in this poly peptide are infertile, every bit their eggs cannot fuse with sperm. The extracellular domain of the α subunit of fertilin contains a hydrophobic region that resembles the fusogenic region of viral fusion proteins, which mediates the fusion of enveloped viruses with the cells that they infect (discussed in Affiliate 13). Synthetic peptides corresponding to this region of the fertilin α concatenation can induce membrane fusion in a examination-tube, consistent with the possibility that fertilin helps to mediate sperm—egg fusion.

Figure 20-33
The fertilin poly peptide in the sperm plasma membrane. The α and β subunits, which are both glycosylated (not shown), are noncovalently associated. Both subunits vest to the ADAM family of proteins, which includes proteins idea to function (more than...)
Male mice that are fertilin-deficient are infertile, and their sperm are eightfold less efficient than normal sperm in binding to the egg plasma membrane but only 50% less efficient in fusing with it. Surprisingly, these defects do not seem to be the master cause of the infertility. The fertilin-deficient sperm are fifty-fifty more dumb in their ability to bind to the zona pellucida and to migrate out of the uterus into the oviduct, where the egg is unremarkably fertilized. Clearly, fertilin'southward roles in fertilization are more complex than originally suspected and are still non completely understood. The finding that fertilin-deficient sperm tin still fertilize eggs in a test tube, albeit inefficiently, suggests that other sperm proteins unremarkably assist to mediate sperm bounden and fusion to the egg plasma membrane.
As the prison cell biology of mammalian fertilization becomes meliorate understood and the molecules that mediate the various steps in the procedure are defined, new strategies for contraception become possible. 1 arroyo currently being investigated, for example, is to immunize males or females with molecules that are required for reproduction in the promise that the antibodies produced will inhibit the activities of these molecules. In addition to the various hormones and hormone receptors involved in reproduction, ZP3 and fertilin might be appropriate target molecules. An alternative approach would be to administrate oligosaccharides or peptides respective to ligands that operate in fertilization, such every bit the postulated integrin-bounden domain of fertilin. Small molecules of this type block fertilization in a test-tube by competing with the normal ligand for its receptor.
The Sperm Provides a Centriole for the Zygote
Once fertilized, the egg is called a zygote. Fertilization is non complete, yet, until the two haploid nuclei (called pronuclei) have come together and combined their chromosomes into a single diploid nucleus. In fertilized mammalian eggs, the two pronuclei do not fuse directly as they do in many other species. They arroyo each other but remain singled-out until after the membrane of each pronucleus has broken down in preparation for the zygote's first mitotic division (Figure xx-34).
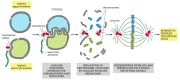
Figure 20-34
The meeting of the sperm and egg pronuclei later on mammalian fertilization. The pronuclei migrate toward the middle of the egg. When they come up together, their nuclear envelopes interdigitate. The centrosome replicates, the nuclear envelopes suspension (more...)
In most animals, including humans, the sperm contributes more than DNA to the zygote. It as well donates a centriole—an organelle that is lacking in unfertilized human eggs. The sperm centriole enters the egg along with the sperm nucleus and tail and a centrosome forms effectually it. In humans, it replicates and helps organize the assembly of the offset mitotic spindle in the zygote (Figure 20-35). This explains why multipolar or extra mitotic spindles form in cases of polyspermy, where several sperm contribute their centrioles to the egg.
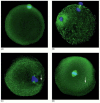
Figure 20-35
Immunofluorescence micrographs of human sperm and egg pronuclei coming together afterward in vitro fertilization. Spindle microtubules are stained in green with anti-tubulin antibodies, and DNA is labeled in blue with a Deoxyribonucleic acid stain. (A) A meiotic spindle in (more...)
Fertilization marks the get-go of one of the almost remarkable phenomena in all of biological science—the process of embryogenesis, in which the zygote develops into a new individual. This is the subject of the side by side chapter.
Summary
Mammalian fertilization begins when the head of a sperm binds in a species-specific manner to the zona pellucida surrounding the egg. This induces the acrosome reaction in the sperm, which releases the contents of its acrosomal vesicle, exposing enzymes that help the sperm to digest its way through the zona to the egg plasma membrane in order to fuse with it. The fusion of the sperm with the egg induces a Catwo+ signal in the egg. The Caii+ signal activates the egg to undergo the cortical reaction, in which cortical granules release their contents, including enzymes that alter the zona pellucida and thereby foreclose the fusion of additional sperm. The Ca2+ signal also triggers the development of the zygote, which begins after sperm and egg haploid pronuclei have come together, and their chromosomes have aligned on a single mitotic spindle, which mediates the first partition of the zygote.

Source: https://www.ncbi.nlm.nih.gov/books/NBK26843/
0 Response to "Describe What Happens to the Chromosomes When an Egg Cell and Sperm Cell Join."
Post a Comment